- International medical Journal & Association for Physicians
АКТУАЛЬНОСТЬ: Растительные лекарственные средства имеют более богатый опыт использования, уходящий глубоко в древность, и продолжают использоваться чаще, чем обычные фармацевтические препараты. Новый сухой экстракт BNO 1011 основан на комбинации из пяти трав, которые используются для лечения острого и хронического риносинусита. Мы оценили фармакологическое действие нового сухого экстракта BNO 1011 на респираторные эпителиальные культуры человека, особенно в отношении транспорта электролита и частоты биения ресничек (СBF).
МЕТОДЫ: Хорошо дифференцированные эпителиальные культуры бронхов человека, выращенные на границе раздела воздух-жидкость, обрабатывали на апикальной или базолатеральной поверхности различными концентрациями сухого экстракта BNO 1011. Изменения трансэпителиального транспорта натрия и хлорида определяли в камерах Уссинга в условиях ограничения напряжения. Изменения в CBF были определены с использованием системы видеоанализа Sissons-Ammons (Ammons Engineering, Mt. Morris, MI).
РЕЗУЛЬТАТЫ: При нанесении на апикальную поверхность сухой экстракт BNO 1011 активировал форсколин-стимулированную секрецию хлорида и цилиарный ритм в зависимости от дозы. Базолатеральное применение сухого экстракта BNO 1011 не изменило измеренных физиологических свойств.
ВЫВОД: Апикальное применение сухого экстракта BNO 1011 стимулирует как секрецию хлоридов, так и CBF и, следовательно, может усиливать мукоцилиарный клиренс.
The novel dry extract BNO 1011 stimulates chloride transport and ciliary beat frequency in human respiratory epithelial cultures
ABSTRACT
Background: Herbal remedies predate written history and continue to be used more frequently than conventional pharmaceutical medications. The novel dry extract BNO 1011 is based on a combination of five herbs that is used to treat acute and chronic rhinosinusitis. We evaluated the pharmacologic effects of the novel dry extract BNO 1011 on human respiratory epithelial cultures specifically addressing electrolyte transport and cilia beat frequency (CBF).
Methods: Well-differentiated human bronchial epithelial cultures grown at an air-liquid interface were treated on the apical or basolateral surface with varying concentrations of dry extract BNO 1011. Changes in transepithelial sodium and chloride transport were determined in Ussing chambers under voltage-clamped conditions. Changes in CBF were determined using the Sissons-Ammons Video Analysis system (Ammons Engineering, Mt. Morris, MI).
Results: When applied to the apical surface, dry extract BNO 1011 activated forskolin-stimulated chloride secretion and ciliary beat in a dose-dependent fashion. Basolateral application of dry extract BNO 1011 did not alter the measured physiological properties.
Conclusion: Apical application of dry extract BNO 1011 stimulates both chloride secretion and CBF and therefore may augment mucociliary clearance.
(Am J Rhinol Allergy 26, 439-443, 2012; doi: 10.2500/ajra.2012.26.3821)
Rhinosinusitis (RS) is a clinically defined syndrome comprising a constellation of symptoms including rhinorrhea, nasal obstruction, facial pressure or pain, headache, and hyposmia. Although the etiology of the disease is not known, a combination of environmental exposures with an underlying genetic predisposition is proposed.1 Although most cases of RS are self-limiting, a subset becomes chronic RS with symptoms lasting >3 months and "characterized by persistent symptomatic, inflammation of the mucosa of the nose and paranasal sinuses."2-3 Impaired function of the sinonasal ciliated mucosa with resultant stasis of sinonasal secretions is a principal element of the pathophysiology of RS4 and is associated with significant local inflammation both in the presence and in the absence of bacterial infection.5 Thus, conventional disease management often includes systemic or topical glucocorticoids to decrease inflammation, systemic or topical antibiotics to combat bacterial infection, and decongestants or mucolytics to restore mucociliary clearance (MCC).
In addition to conventional therapies, >40% of patients with RS use complementary and alternative therapies with over one-quarter using herbal preparations.6 One such product, Sinupret (Bionorica SE, Neu- markt, Germany), is a trademarked medicinal product consisting of five herbs: gentian root (Gentianae radix), primrose flowers (Primulae flos), elder flowers (Sambuci flos), common sorrel herb (Rumicis herba), and vervain herb (Verbenae herba) in the fixed ratio of 1:3:3: 3:3. Three randomized trials investigated the effect of this preparation as an adjunct treatment for acute RS in patients being treated with antibiotics.7 In two of the trials, patients taking oral Sinupret showed statistically significant improvement in outcomes, and one study showed a trend toward improvement but did not reach significance. These clinical results prompted investigators to hypothesize that Sinupret might have direct effects on the respiratory epithelium. In mouse nasal epithelial cells both in vivo and in vitro, Sinupret caused a dose-dependent increase in Cl- secretion, leading the investigators to conclude that Sinupret stimulates airway surface liquid hydration via a cystic fibrosis transmembrane conductance regulator (CFTR)- dependent mechanism.8 Similar results were observed with another flavonoid compound, quercetin, in human respiratory epithelia.9 Because there are both similarities and differences between murine and human airway epithelial cells and because Sinupret showed positive effects on murine epithelia, we tested the hypothesis that the novel Sinupret dry extract would alter ion transport and ciliary beating in a model of human airway epithelia. We used commercially available human bronchial epithelial (HBE) cells grown at an air-liquid interface (ALI) to evaluate the effect of Sinupret dry extract (BNO 1011) on human respiratory ion transport and ciliary beating properties.
METHODS Cell Culture
HBE cells from normal donors were purchased from Lonza (Walk- ersville, MD) and expanded in Bronchial Epithelial Cell Growth Medium (BEGM; Lonza) as previously described.10-13 A total of six tissue donors were used in these experiments.
Materials
Sinupret dry extract (BNO 1011) is a native dry extract. The dry extract BNO 1011 was prepared from the Sinupret drug mixture composed of Gentianae radix, Primulae flos, Sambuci flos, Rumicis herba, and Verbenae herba in the fixed ratio of 1:3:3:3:3 with 59% ethanol as extracting agent (v/v), resulting in a final drug/extract ratio of 5.9:1. The quality of herbal drugs (starting material) is specified according to the relevant European Medicines Agency guide lines for herbal medicinal products. BNO 1011 was manufactured in a validated production process according to good manufacturing practice. Comprehensive specifications and standardized production processes guarantee high batch-to-batch consistency. For Ussing chamber and ciliary beat experiments, BNO 1011 was prepared per the following instructions: 250 mg of BNO 1011 was dissolved in 1 mL of 50% ethanol and bath sonicated at 35 kHz for 30 minutes, followed by centrifugation at 3000 X g for 10 minutes at room temperature with careful aspiration of the supernatant (stock of 250 mg/mL). Final solutions were then diluted in either physiological bath solution (for apical application) or BEGM (for basolateral application) to the indicated concentrations. Amiloride (Sigma, St. Louis, MO) was dissolved in distilled, deionized water as 1000 X stock. Forskolin and inhibitor of CFTR (CFTRinh-172; Calbiochem, EMD, San Diego, CA) were dissolved in DMSO as 10- and 30-mM stock solutions, respectively.
Electrophysiology
Short-Circuit Current Measurements. The Ussing chamber bath solution containedl20 mM of NaCl, 25 mM of NaHC03, 3.3 mM of KH2P04, 0.8 mM of K2HP04,1.2 mM of MgCl2,1.2 mM of CaCl2, and 10 mM of glucose. Short-circuit currents (Isc) were measured as previously described.12-13 Briefly, transwell inserts (Corning Life Sciences, Corning, NY) were placed inside a modified, vertical Ussing chamber (Physiologic Instruments, San Diego, CA) housed in a 37°C heated manifold, and the monolayers were continuously voltage clamped to 0 mV using automatic voltage clamps (VCC 600; Physiologic Instruments) after adjusting for fluid resistance. Transwell filters were mounted in bath solution warmed to 37°C, and the solution was continuously infused with 95% Oz-5% COz. Isc was digitized at one sample every 10 seconds and data were stored on a computer hard drive using Acquire & Analyze software build 2.2 (Physiologic Instruments). Transepithelial resistance was calculated by computer program, which placed a 640-ms, bipolar 10-mV potential across the monolayer in between each data point and calculated resistance by Ohm's law (transepithelial resistance = ДУ/ДI). By convention, a positive deflection in 1^ represents anion secretion or cation absorption. Experiments were repeated on cells from a minimum of three different lots of HBE cells.
Ciliary Beat Frequency
Ciliary beat frequency (CBF) measurement was performed as previously described.14-15 Images of beating cilia on the HBE ALI cultures were visualized using a 63X objective on an inverted scope (Leica Microsystems, Inc., Bannockburn, IL). Image data were captured using a model A602f-2 Basler area scan high-speed monochromatic digital video camera (Basler AG, Ahrensburg, Germany) at a sampling rate of 100 frames/s with a resolution of 640 X 480 pixels. The video images were analyzed using the Sisson-Ammons Video Analysis (SAVA) system version 2.1.15 (Ammons Engineering).16 For each experiment, a large area of beating cilia on the ALI cultures was detected with the inverted microscope. The digital image signal was then routed from the camera directly into a digital image acquisition board (National Instruments, Austin, TX) within a Dell XPS 710 Workstation (Roundrock, TX) running Windows XP Professional operating system. Images were captured at 100 frames/s, compressed, and stored to disk. Files were reloaded and analyzed with virtual instrumentation software highly customized to perform CBF analysis. All of the recordings in the present experiments were made at 630 X magnification.
All experiments were performed at an ambient temperature of 26°C and before all data acquisition, 40 mL room temperature sterile PBS was added to the apical surface of the ALI cultures and allowed to equilibrate to 26°C for 10 minutes.17'18 To obtain basal CBF for each culture, 10 videos (2 seconds at a frame rate of 100 frames/s) were randomly obtained at different locations of the ALI.
To determine the effect of BNO 1011 on basal CBF of the HBE ALI cultures, after temperature equilibration, cultures were treated either on the apical surface with a 10 X solution or in the basolateral side with a 100X solution to obtain the final designated concentrations (0.1, 1, and 2.5 mg/mL). CBF was then obtained as described previously at 10, 30, 60, and 120 minutes and 24 hours.
Data Presentation and Statistical Analysis
For electrophysiological experiments we report Igc data as mean Igc ± standard error for the total number of inserts tested. Change in 1^ was calculated by subtracting the 1^ just before addition of a compound from the Igc after stabilization after addition of that compound. Comparisons were made with ANOVA followed by posttests to determine significance between individual groups. All statistical comparisons were made using Prism 5 (GraphPad Software, San Diego, CA). Significance was defined as a value of p s 0.05. For CBF experiments statistical analysis of the arithmetic means derived from each culture was performed using two-tailed paired t-tests.
RESULTS
We first evaluated the pharmacologic effects of BNO 1011 on trans-epithelial electrolyte transport. BNO 1011 was added in increasing concentrations to the basolateral surface of HBE cells mounted in Ussing chambers before the sequential addition of amiloride and forskolin. There was no significant difference in baseline Isc, amiloride-sensitive Isc, or forskolin-stimulated Isc after basolateral addition of BNO 1011 (Fig. 1 A). In a second set of experiments, BNO 1011 was added to the basolateral bath of the Ussing chamber after addition of amiloride to the apical bath to isolate the potential effects of BNO 1011 on forskolin-stimulated Isc. Again, there was no difference between BNO 1011-treated cells and vehicle controls (Fig. 1 B). Analysis of basolateral application of BNO 1011 on ciliary activity yielded similar results with no change in CBF at multiple time points and concentrations compared with the vehicle-treated cultures (Fig. 1 C).
Prior work in mouse epithelial cells indicated that simultaneous application of Sinupret to the apical and basolateral surface of polarized respiratory epithelial cultures induced a chloride efflux.8 Therefore, to investigate if apical addition of BNO 1011 would have effects at the apical membrane of human epithelial cells, we repeated our experiments with apical application of BNO 1011. When BNO 1011 was added in increasing amounts to the apical bath before sequential addition of amiloride and forskolin, there was a trend toward increased baseline Isc, decreased amiloride-sensitive Isc, and decreased forskolin-stimulated Isc (Fig. 2 A), suggesting that BNO 1011 might activate anion secretion through a cAMP-mediated pathway. Consistent with this observation, when BNO 1011 was added after the addition of amiloride, there was a significant dose-dependent increase in Isc compared with vehicle controls that was paralleled by a decrease in the change in Isc after addition of forskolin (Fig. 2, В and C), suggesting that the two stimuli, BNO 1011 and forskolin, act through the same pathway. Similarly, subsequent to apical application of BNO 1011, a dose-dependent significant increase in CBF was evident that lasted for up to 2 hours at the highest concentrations tested (Fig. 2 D).
DISCUSSION
Mucus overproduction and mucus stasis are common sequelae of respiratory infections and inflammatory processes that may result in upper and lower respiratory tract morbidities including nasal congestion, rhinorrhea, facial pain and pressure, cough, wheezing, and chest pain. Because of the negative impact that these symptoms may have on quality of life, 19 multiple pharmaceutical and herbal medicines have been used in an attempt to alleviate them. Many of these therapies are available over-the-counter, and, to date, the effects of these remedies on the physiology of respiratory epithelial cells is neither well studied nor well understood. The commercially available oral remedy Sinupret has previously been studied in mouse models but not in cells of human origin. Our data presented here suggest that BNO 1011, a novel dry extract prepared from the herbal drug mixture contained in Sinupret promotes anion secretion and ciliary beating in normal HBE cells when applied to the apical surface but not the basolateral surface.
Specifically, we showed that BNO 1011 caused a dose-dependent, forskolin-independent increase in Isc in well-differentiated HBE cells. Moreover, after application of BNO 1011, the change in Isc after forskolin was decreased, but the total Isc inhibited by CFTRinh-172, a relatively specific inhibitor of CFTR,20 was not significantly decreased, suggesting that BNO 1011 acts by increasing CFTR-mediated ion transport. Finally, addition of BNO 1011 after doses of forskolin that maximally stimulate CFTR activity (2 /nM) did not result in further increases in Isc. Forskolin, a diterpene compound, activates membrane adenylyl cyclase thereby raising cytoplasmic cAMP. cAMP, in turn, activates protein kinase A that phosphorylates, and thereby activates, the CFTR. Therefore, forskolin stimulation of respiratory epithelial cells in the presence of amiloride and a favorable electrochemical driving force results primarily in chloride and bicarbonate secretion, which are reflected as increased Isc. BNO 1011 similarly increased Isc. Taken together, these data suggest that BNO 1011 stimulates CFTR-mediated anion secretion. In contrast to its effects on forskolin-stimulated anion secretion, BNO 1011 did not appear to have a significant effect on amiloride-sensitive Isc, a measure of epithelial Na+ channel-mediated sodium absorption.
These data agree with previously published data in mouse models.8 They are also consistent with the findings of investigators who studied the flavonoid compound, quercetin, that activates CFTR in human airway epithelial cells.21 Quercetin-induced CFTR activation was in-dependent of CFTR R-domain phosphorylation, suggesting that it may directly activate CFTR rather than activating CFTR through the cAMP-protein kinase A second messenger system, which would result in R-domain phosphorylation.22 The complex flavonoid and phytochemical mixture BNO 1011 may work in a similar manner, although our data do not directly address this possibility.
There is a close relationship between transepithelial ion transport, homeostasis of airway surface liquid, ciliary beating, and MCC. For example, in patients with CF, a multisystem disease of altered ion transport, absence of CFTR in respiratory epithelial cells leads to impaired MCC throughout the respiratory tract.23 The precise etiology of impaired MCC in CF is not fully delineated, although there are at least two nonmutually exclusive hypotheses. First, lack of CFTR leads to excess sodium (Na+) absorption through the epithelial Na+ channel in airway epithelial cells.24-25 This Na+ hyperabsorption leads to airway surface liquid (ASL) dehydration, which impairs MCC because adequate ASL volume is required for proper ciliary beating.23 Second, lack of CFTR results in chloride and bicarbonate hypose- cretion from airway epithelial cells26 and submucosal glands,27 resulting in inadequate fluid secretion and therefore decreased ASL volume and inadequate MCC. In patients with primary ciliary dysfunction there is impaired MCC throughout the respiratory tract, which is independent of electrolyte transport.28 Finally, in multiple acquired respiratory infections, such as chronic RS, there is evidence suggesting both impaired electrolyte transport29 as well as ciliary dysfunction30 contribute to impaired MCC.
These observations logically lead to the hypothesis that increasing epithelial anion transport, ciliary beating, or both in the setting of impaired MCC may be of therapeutic benefit. BNO 1011 applied to the apical membrane, but not the basolateral membrane, of human airway epithelial cells acutely increased both anion secretion and ciliary beating, which together would be expected to increase MCC. Were BNO 1011 to be administered orally, one might expect it to have an acute effect on MCC only if the active compound is secreted or passively moves onto the surface of the respiratory epithelium. Our studies suggest that BNO 1011 does not rapidly cross the epithelium as modeled in our culture system, but these differentiated cells may not completely reflect the situation in vivo, where active secretion of BNO 1011 into the pericellular or mucus layer above the epithelium could lead to the compound activating epithelial chloride secretion. Furthermore, we cannot completely exclude the possibility that BNO 1011 acts intracellularly and that application to the apical membrane allows the active compound to diffuse into the cell to a greater degree than does application to the basolateral membrane. Additional studies are necessary to determine the exact mechanisms by which BNO 1011 activates anion secretion and ciliary beating, although it is likely that the mechanism of action is similar to other flavonoid compounds.9'21 Therefore, if further investigation does not reveal movement of secretion of BNO 1011 across the epithelium, a further consideration might be the preparation of a topical application so that the apical surface of the epithelium is directly contacted by the medicine. Furthermore, it should be noted that these studies were performed using a solution prepared from the dried extract rather than direct application of the dried extract itself. We do not expect that the method of preparation had an impact on the results because vehicle control experiments were performed with equal dilutions of the stock 50% ethanol and did not show the same activation of chloride secretion. Additionally, no attempt was made to fractionate the dried extract, and, therefore, we cannot speculate as to which of the compounds are required to activate chloride secretion. Rather, we can conclude that together the components are sufficient to activate chloride secretion.
In summary, our data presented here are consistent with those of other investigators who studied Sinupret in murine systems and who studied the flavonoid quercetin in human airway epithelial cells. Taken together, these findings suggest that BNO 1011 may in fact have pharmacologic effects that promote MCC (increased anion secretion and ciliary beating), and additional studies to determine the efficacy of these compounds in diseases of altered MCC may be warranted.
James L. Kreindler, M.D., Bei Chen, M.D., Yael Kreitman, M.D.,
Jennifer Kofonow, M.S., Kelly M. Adams, M.D.,
and Noam A. Cohen, M.D., Ph.D.
From the 1Children's Hospital of Philadelphia and Department of Pediatrics, University of Pennsylvania School of Medicine, Philadelphia, Pennsylvania,? Department of Otorhinolaryngology, Head and Neck Surgery, University of Pennsylvania, Philadelphia, Pennsylvania, 3TempIe University School of Medicine, Philadelphia, Pennsylvania, and ^Philadelphia Veterans Affairs Medical Center, Surgical Services, Philadelphia, Pennsylvania
Funded by Bionorica SE, Neumarkt, Germany. The company had no role in collecting or analyzing data for inclusion in this article but did participate in its generation. Evaluation of raw data and data interpretation were performed independently by the investigators
The authors have no conflicts of interest to declare pertaining to this article В Chen and Y Kreitman contributed equally to this article
Address correspondence and reprint requests to Noam A. Cohen, M.D., Ph.D., Department of Otorhinolaryngology - Head and Neck Surgery, Ravdin Building 5th floor, 3400 Spruce Street, Philadelphia, PA 19104 E-mail address: cohenn@uphs.upenn.edu Copyright © 2012, OceanSide Publications, Inc., U.S.A.
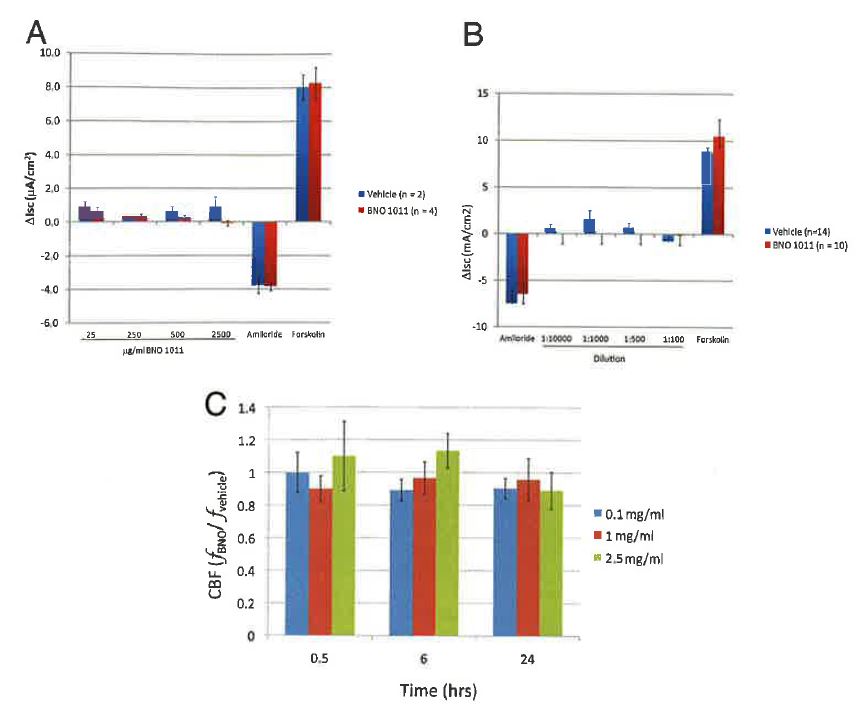
Figure 1. Application o/BNO 1011 to the basolateral (serosal) surface of the cells. (A) BNO 1011 (stock solution of 250 mg/mL) was added sequentially in increasing amounts to the serosal bath of human bronchial epithelial (HBE) cells mounted in Ussing chambers (final concentrations: 25, 250, 500, and 2500 p.g/mL). After the last addition, amiloride (10 pM) was added to the apical (mucosal) bath and forskolin (2 pM) to both apical and basolateral batlis to inhibit sodium absorption and stimulate chloride secretion, respectively. (B) Amiloride (10 pM) was added to the apical solution to block sodium absorption. Then, BNO 11011 was added sequentially in increasing amounts to the basolateral bath (final concentrations: 25, 250, 500, and 2500 pg/mL). After the last addition, forskolin (2 pM) was added to both surfaces to stimulate chloride secretion. (C) Addition of BNO 1011 to the basolateral surface does not affect ciliary beat frequency (CBF; n = two patients, three cultures from each patient/condition).
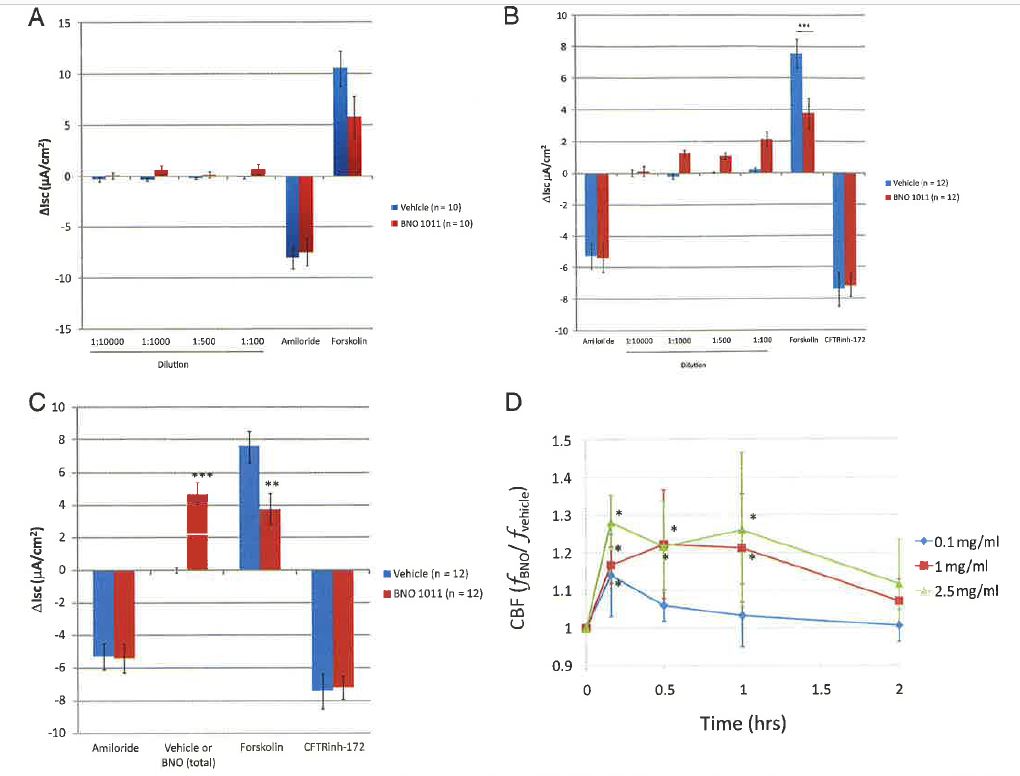
Figure 2. Application of BNO 1011 to the apical (mucosal) surface of the cells. (A) BNO 1011 (stock solution of 250 mg/mL) was added sequentially in increasing amounts to the apical bath of human bronchial epithelial (HBE) cells mounted in Ussing chambers (final concentration: 25, 250, 500, and 2500 p-g/mL). After the Inst addition, amiloride was added to the apical (mucosal) bath and forskolin to both apical and basolateral baths to inhibit sodium absorption and stimulate chloride secretion, respectively. (B) Amiloride (10 p.M) was added to the apical solution to block sodium absorption followed by addition of BNO 1011 in sequentially increasing concentrations (final concentrations: 25, 250, 500, and 2500 jxg/mL). After the last addition, forskolin (2 p,M) was added to both baths to stimulate chloride secretion. (C) Cumulative change in l^from panel B. (D) Addition of BNO 1011 to the apical surface stimulates sustained elevation in ciliary beat frequency (CBF) in a dose-dependent manner (n = three patients, three cultures from each patient/condition; *p < 0.05, **p < 0.01, and ***p < 0.001 versus vehicle).
REFERENCES
1. Kern RC, Conley DB, Walsh W, et al. Perspectives on the etiology of chronic rhinosinusitis: An immune barrier hypothesis. Am I Rhinol 22:549-559, 2008.
2. Fokkens WJ, Lund VJ, Mullol J, et al. EPOS 2012: European position paper on rhinosinusitis and nasal polyps 2012. A summary for oto- rhinolaryngologists. Rhinology 50:1-12, 2012.
3. Meltzer EO, and Hamilos DL. Rhinosinusitis diagnosis and manage-ment for the clinician: A synopsis of recent consensus guidelines. Mayo Clin Proc 86:427-443, 2011.
4. Gudis D, Zhao KQ, and Cohen NA. Acquired cilia dysfunction in chronic rhinosinusitis. Am J Rhinol Allergy 26:1-6, 2012.
5. Hsu J, and Peters AT. Pathophysiology of chronic rhinosinusitis with nasal polyp. Am J Rhinol Allergy 25:285-290, 2011.
6. Blanc PD, Trupin L, Earnest G, et al. Alternative therapies among adults with a reported diagnosis of asthma or rhinosinusitis: Data from a population-based survey. Chest 120:1461-1467, 2001.
7. Guo R, Canter PH, and Ernst E. Herbal medicines for the treatment of rhinosinusitis: A systematic review. Otolaryngol Head Neck Surg 135:496-506, 2006.
8. Virgin F, Zhang S, Schuster D, et al. The bioflavonoid compound, sinupret, stimulates transepithelial chloride transport in vitro and in vivo. Laryngoscope 120:1051-1056, 2010.
9. Zhang S, Smith N, Schuster D, et al. Quercetin increases cystic fibrosis transmembrane conductance regulator-mediated chloride transport and ciliary beat frequency: Therapeutic implications for chronic rhi- nosinusitis. Am J Rhinol Allergy 25:307-312, 2011.
10. Kreindler JL, Bertrand CA, Lee RJ, et al. Interleukin-17A induces bicarbonate secretion in normal human bronchial epithelial cells. Am J Physiol Lung Cell Mol Physiol 296:L257-L266, 2009.
11. Kreindler JL, Jackson AD, Kemp PA, et al. Inhibition of chloride secretion in human bronchial epithelial cells by cigarette smoke ex-tract. Am J Physiol Lung Cell Mol Physiol 288:L894-L902, 2005.
12. Savitski AN, Mesaros C, Blair IA, et al. Secondhand smoke inhibits both Cl- and K+ conductances in normal human bronchial epithelial cells. Respir Res 10:120, 2009.
13. Zhao KQ, Xiong G, Wilber M, et al. A role for two-pore K+ channels in modulating Na+ absorption and CL secretion in normal human bronchial epithelial cells. Am J Physiol Lung Cell Mol Physiol 302: L4-L12, 2012.
14. Woodworth BA, Zhang S, Tamashiro E, et al. Zinc increases ciliary beat frequency in a calcium dependent manner. Am J Rhinol Allergy 24:6-10, 2010.
15. Zhao KQ, Goldstein N, Yang H, et al. Inherent differences in nasal and tracheal ciliary function in response to Pseiidomoms aeruginosa challenge. Am J Rhinol Allergy 25:209-213, 2011.
16. Sisson JH, Stoner JA, Ammons BA, and Wyatt ТА. All-digital image capture and whole-field analysis of ciliary beat frequency. J Microsc 211:103-111, 2003.
17. Schipor I, Palmer JN, Cohen AS, and Cohen NA. Quantification of ciliary beat frequency in sinonasal epithelial cells using differential interference contrast microscopy and high-speed digital video imag-ing. Am J Rhinol 20:124-127, 2006.
18. Zhao KQ, Cowan AT, Lee RJ, et al. Molecular modulation of airway epithelial ciUary response to sneezing. Faseb J 26:3178-3187, 2012.
19. Gliklich RE, and Metson R. The health impact of chronic sinusitis in patients seeking otolaryngologic care. Otolaryngol Head Neck Surg 113:104-109, 1995.
20. Thiagarajah JR, Song Y, Haggie PM, and Verkman AS. A small molecule CFTR inhibitor produces cystic fibrosis-like submucosal gland fluid secretions in normal airways. Faseb J 18:875-877, 2004.
21. Pyle LC, Fulton JC, Sloane PA, et al. Activation of the cystic fibrosis transmembrane conductance regulator by the flavonoid quercetin: Potential use as a biomarker of DeltaF508 cystic fibrosis transmem-brane conductance regulator rescue. Am J Respir Cell Mol Biol 43: 607-616, 2010.
22. Cheng SH, Rich DP, Marshall J, et al. Phosphorylation of the R domain by cAMP-dependent protein kinase regulates the CFTR chlo¬ride channel. Cell 66:1027-1036, 1991.
23. Boucher RC. Evidence for airway surface dehydration as the initiat-ing event in CF airway disease. J Intern Med 261:5-16, 2007.
24. Gentzsch M, Dang H, Dang Y, et al. The cystic fibrosis transmem-brane conductance regulator impedes proteolytic stimulation of the epithelial Na+ channel. J Biol Chem 285:32227-32232, 2010.
25. Trout L, King M, Feng W, et al. Inhibition of airway liquid secretion and its effect on the physical properties of airway mucus. Am J Physiol 274:L258-L263, 1998.
26. Smith JJ, and Welsh MJ. cAMP stimulates bicarbonate secretion across normal, but not cystic fibrosis airway epithelia. J Clin Invest 89:1148-1153, 1992.
27. Joo NS, Irokawa T, Robbins RC, and Wine JJ. Hyposecretion, not hyperabsorption, is the basic defect of cystic fibrosis airway glands. J Biol Chem 281:7392-7398, 2006.
28. Moller W, Haussinger K, Ziegler-Heitbrock L, and Heyder J. Muco-ciliary and long-term particle clearance in airways of patients with immotile cilia. Respir Res 7:10, 2006.
29. Dejima K, Randell SH, Stutts MJ, et al. Potential role of abnormal ion transport in the pathogenesis of chronic sinusitis. Arch Otolaryngol Head Neck Surg 132:1352-1362, 2006.
30. Chen B, Shaari J, Claire SE, et al. Altered sinonasal ciliary dynamics in chronic rhinosinusitis. Am J Rhinol 20:325-329, 2006.
30.01.2020 5421